
The goal of the research performed in this laboratory is to investigate the neural basis of perception. More specifically, we want to understand how cortical microcircuits process sensory information leading to behavioral outcomes.
We use optogenetic and pharmacological approaches in our research to turn neurons 'ON' and 'OFF'. In doing so, we can observe and manipulate specific subsets of neurons at multiple levels: in slice preparations in vitro, and in anesthetized or awake, behaving animals in vivo. This approach allows us to quantitatively determine how specific subsets of neurons contribute to sensory processing and behavior.
By complementing our in vivo work with synaptic connectivity and network dynamics analysis in vitro, we achieve a more complete understanding about how neural circuits in our brain support sensation, action, and cognition.
Current Project
Callosal Projections Drive Neuronal-Specific Responses in the Mouse Auditory Cortex
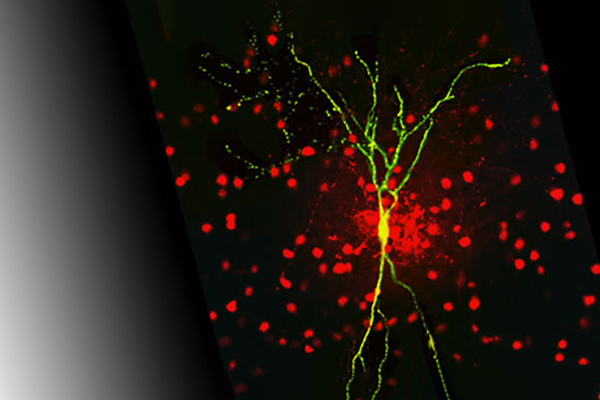
Right: A layer 2/3 fast-spiking interneuron of the auditory cortex
Cortical microcircuits process sensory information to drive behavior. Deciphering how populations of neurons encode information, generate perceptions, and execute behavioral decisions requires working at both the cellular and system levels.
Publications
Rock C, Zurita H, Wilson C, Apicella Aj (2016). An inhibitory corticostriatal pathway. Elife May 9;5.
Rock C, Apicella Aj (2015). Callosal Projections Drive Neuronal-Specific Responses in the Mouse Auditory Cortex. Journal of Neuroscience 35(17):6703-6713.
Apicella Aj, Wickersham IR, Seung HS,Shepherd GMG (2012). Laminarly Orthogonal Excitation of Fast-Spiking and Low-Threshold-Spiking Interneurons in Mouse Motor Cortex. Journal of Neuroscience 32(20):7021-33.
Apicella Aj, Qi Yuan, Scanziani M and Isaacson JS (2010). Pyramidal Cells in Piriform Cortex Receive Convergent Input from Distinct Olfactory Bulb Glomeruli. Journal of Neuroscience 30(42):14255-14260.
Li B, Devidze N, Barengolts D, Prostak N, Sphicas E, Apicella Aj, Malinow R and Emamian ES (2009). NMDA Receptor Phosphorylation at a Site Affected in Schizophrenia Controls Synaptic and Behavioral Plasticity. Journal of Neuroscience 29(38):11965-11972.
Apicella Aj, Wang Y, Lee SK, Eczurra M, Slone D, Goldmit M, Schafer WR, Shaham S, Driscoll M, Bianchi L (2008). A glial DEG/ENaC channel functions with neuronal channel DEG-1 to mediate specific sensory functions in C. elegans. EMBO Journal 27:2388-2399.
Apicella Aj, Hilliard MA, Kerr R, Suzuki H, Bazzicalupo P and Schafer WR (2005). In vivo imaging of C. elegans ASH neurons: cellular response and adaptation to chemical repellents. EMBO Journal 24:63-72.
Kahn-Kirby AH, Dantzker JLM, Apicella Aj, Schafer WR, Browse J, Bargmann CI, Watts JL (2004). Specific Polyunsaturated Fatty Acids Drive TRPV-Dependent Sensory Signaling In Vivo. Cell 119(6):889-900.
Fukuto HS, Ferkey DM, Apicella Aj , Lans H, Sharmeen T, Chen W, Lefkowitz RJ, Jansen G, Schafer WR and Hart AC. (2004). G Protein-Coupled Receptor Kinase Function Is Essential for Chemosensation in C. elegans. Neuron 42(4):581-593.
Ursini MV, Gaetaniello L, Ambrosio R, Matrecano E, Apicella Aj, Salerno MC. and Pignata C. (2002). Atypical X-linked SCID phenotype associated with growth hormone hyporesponsiveness. Clinical and Experimental Immunology 129:502-9.
